Squishy things, or a new post not about education
My first semester of University involved taking BIOSCI 103: Comparative Animal Biology. It was a great course, and really piqued my interest in pursuing an animal-based biology degree, rather than a biochemistry one, which had been my initial intention. Hell, my initial intial intention had been to do law (until I worked for lawyers), so it's not like changing my mind as often as I change my underwear isn't particularly out of the ordinary...
Nonetheless, I digress. The course itself contained laboratories which essentially involved hacking all manner of animals up and looking at their innards. My initial uneasiness wore off in about 3 minutes into the first lab, as I realised how cool chopping animals up can be. Ok, sure, it's probably not very ethical to assign one rat per person, rats that had been specifically bred to be slaughtered, congealed blood staining their nostrils from the poison-induced respiratory haemorrhaging that ultimately led to their demise. But hey. I had just bought my new leather bound dissection kit, and like I wasn't going to let that go to waste, right?
Oh dear. Another digression.
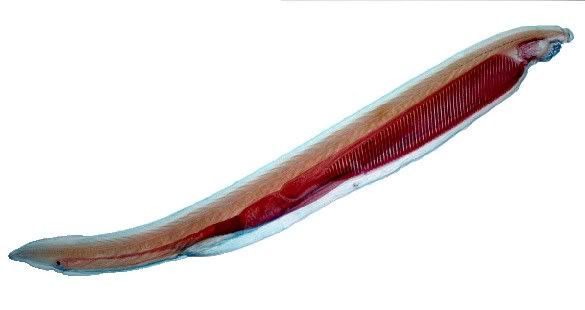
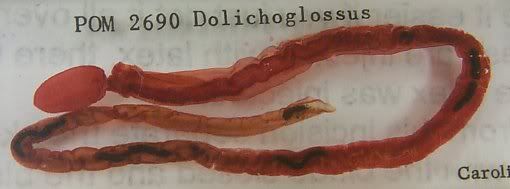
ANYway. One of our labs involved observation of Amphioxus, or the lancelet 'worm', under the microscope. These critters are only very small, about 2-3cm maximum in length, and look a lot like anchovies. Amphioxus, otherwise called Branchiostoma has long been lauded as a key model species for understanding the development of the vertebrates from from an invertebrate world sometime during the Ordovician period of the Paleozoic era, some 470 million years ago.
Our little corner of the tree of life is called the Deuterostomia (which means "second mouth"), as opposed to Protostomia (which means "first mouth"). These names refer to the period of development where the gut forms: in protostomes, the first opening eventually forms the mouth, whereas in deuterostomes (us), the first opening forms the anus, and the second opening forms the mouth - hence 'second mouth'. Comprising this set are the echinoderms (star fish, kina et cetera), the hemichordates (a random and obscure group of worms) and the 'chordates'. The chordates themselves split into the urochordates - things like sea squirts and other tunicates, the cephalochordates, where our little lancelet friend fits in, and the vertebrates - us.
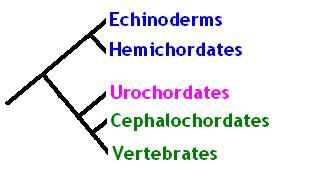
The textbook view of these groups holds that echinoderms are the most 'primitive', or basal member of the deuterostomes. This is mainly due to the fact that they are unlike any other deuterostome - they are the odd ones out in terms of their symmetry, their locomotion, internal organisation et cetera. The next group up are the hemichordates, so called because they do have some of the characteristics which are normally associated with the chordates, in particular the presence of first signs of gill (pharyngeal) slits. The classical view argues that the urochordates (tunicates) are the most basal member of the chordate clade - they possess a hollow 'notochord' and dorsal nerve cord, but don't exhibit the body complexity, particularly body segmentation, seen in the remaining cephalochordates (lancelets) and vertebrates, which together comprise the Euchordata or 'true chordates'. This classical view is shown in the image below.

[Click to enlarge]
All of this is based on an apparent increase in complexity throughout the lineages - almost as if the principles behind that iconic 'ape to man' idea had been transplanted to explain the development from 'squishy' to 'us'. Those more like us are more advanced, those more squishy are, well, primitive. Of course, we're only dealing here with complexity in a morphological sense - the appearance of new and improved features on organisms over time. If we were to look at biochemical or intracellular physiological complexity, the earliest forms of life (bacteria) would kick our proverbial arse. This idea of a supposed correlation between being 'complex' and being 'advanced' isn't completely arrogant or conceited. The earliest forms of animal life at least were simple and squishy, and there is, overall, a general trend towards "complexity". But many squishy lineages developed incredible complexity - you only need to look at the cephalopods (octopuses, squid and cuttlefish) who are most closely related to snails and slugs, as well as the more 'complex' forms that have rediscovered the joys of a simple squishy lifestyle.
This view on the deuterostome phylogeny has been challenged before, with some success. However, the most significant recent change to the phylogeny really only focussed on the origins of the hemichordates (shown below), shifting them from being basal to the chordates, to being a more closely related sister taxon of the echinoderms. As you can see, that still maintains that nice, convenient transition from the more primitive and squishy sea squirts, through Amphioxus, up to us and the other vertebrates.
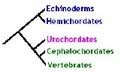
[Click to enlarge]
Not anymore.
A paper published in this last week's Nature throws some quite serious spanners into the classical works of deuterostome phylogeny. The researchers, Delsuc et al. (Delsuc interestingly held a post-doctoral position in Massey Univerity's Allan wilson Centre for Ecology and Evolution) used 146 common genes from 14 different deuterostomes to really hammer out the relationships between the groups. The results below are quite profound (ignore the black bits, just focus on the bottom coloured bits).

[Click to enlarge]
Essentially this provides two fundamental shifts:
Firstly, the urochordates are placed as the sister group to the Vertebrates, supporting a previously hypothesised clade called Olfactores. What this phylogeny implies is that we are more closely related to the sea squirt we are desperately trying to get rid of in Auckland Harbour than we are to Amphioxus. Now, this may not sound very important to some of you, but for anyone who has done even some basic evolutionary study, it means everything that we assume about the evolution and appearance of the vertebtrates, this gradual appearance of more 'vertebrate' features via the cephalochordates could quite possibly be absolute shite. Ipso facto, our (perhaps conceited) grouping "Euchordata" or "True chordates" could also be absolute shite.
Secondly, The cephalochordates are not only not directly basal to the vertebrates, they are actually placed as the sister group to the echinoderms in their own clade. Although the authors point out that this particular grouping doesn't have the statistical robustness as the urochordate-vertebrate grouping, it is nonetheless a rational interpretation of their data. This completely disrupts the "Chordata" group. From a systematics point of view, a lineage name applies to a common ancestor and all it's living and non-living descendants. Given that the chordates are now distributed all over the show (that is, they are polyphyletic) the term "chordate" may not, evolutionarily speaking, mean a hell of a lot. Unless of course we lowered the bar, if you will, and classified echinoderms (and by default hemichordates) as chordates. This naturally, is not likely to happen, because then basically we would be renaming all of the living deuterostomes as chordates. And that just won't do. No it won't. These two shifts can be seen in the simplified cladogram below.

[Click to enlarge]
This is the beauty of the tension that often arises between traditional morphological cladistics, or grouping organism relatedness based on overall similarity of body form and the sharing of structures, and that of molecular systematics - using genes to work out the same thing. Often they do come out with the same answer, but many times they don't. It's incredibly exciting when a study like this comes out, and challenges our assumptions about our 'knowledge' of morphological systems.
Perhaps more exciting than the actual repositioning of taxa within the deuterostome phylogeny is the implication for what this repositioning means for the evolutionary history of the deuterostomes themselves. The classical view suggests a steady aquisition and appearance of more 'vertebrate' like features through the lineages, with the cephalochordates representing the penultimate innovation before the appearance of the first vertebrate fish about 470 million years ago. Conversely, the echinoderms are seen (along with their sister group the hemichordates) as some bygone era of deuterstome evolution, a weird and wacky cousin that doesn't quite fit. Similarly, the urochordates, with their sessile habit and (apparently) simple body plan are seen as the primitive precursors to the euchordates.
According to the new study, however, both of these taxa can be viewed as highly advanced and derived lineages, which extensively modified the ancestral deuterostome body plan to become the forms that we see today. In the echinoderms an internal calcitic skeleton developed, along with radial symmetry and a particularly unique mode of locomotion, in the urochordates, the complete loss of the notochord in at least one sublineage, unique genetic simplicity and a lazy, squishy way of life.
And what of that ancestral body plan? The researchers suggest that the last common ancestor of the living deuterostomes may be far closer to Amphioxus than previously imagined - free living, bilateral symmetry, well developed metamerism (segmentation), distinct gill slits, and a pretty well organised brain and dorsal central nervous system. Essentially, all of these features that we assumed appeared just before we did may actually be the most ancestral, the most basic condition, Further those forms we assumed to be 'primitive' and distantly related are in fact extreme modifications of that basic body plan, bizarre yet equisite examples of evolution and adaptation, and in the case of the urochordates, closer to us in evolutionary time than we had ever imagined.
Nonetheless, I digress. The course itself contained laboratories which essentially involved hacking all manner of animals up and looking at their innards. My initial uneasiness wore off in about 3 minutes into the first lab, as I realised how cool chopping animals up can be. Ok, sure, it's probably not very ethical to assign one rat per person, rats that had been specifically bred to be slaughtered, congealed blood staining their nostrils from the poison-induced respiratory haemorrhaging that ultimately led to their demise. But hey. I had just bought my new leather bound dissection kit, and like I wasn't going to let that go to waste, right?
{kind=link}
Oh dear. Another digression.
ANYway. One of our labs involved observation of Amphioxus, or the lancelet 'worm', under the microscope. These critters are only very small, about 2-3cm maximum in length, and look a lot like anchovies. Amphioxus, otherwise called Branchiostoma has long been lauded as a key model species for understanding the development of the vertebrates from from an invertebrate world sometime during the Ordovician period of the Paleozoic era, some 470 million years ago.
{kind=link}
Our little corner of the tree of life is called the Deuterostomia (which means "second mouth"), as opposed to Protostomia (which means "first mouth"). These names refer to the period of development where the gut forms: in protostomes, the first opening eventually forms the mouth, whereas in deuterostomes (us), the first opening forms the anus, and the second opening forms the mouth - hence 'second mouth'. Comprising this set are the echinoderms (star fish, kina et cetera), the hemichordates (a random and obscure group of worms) and the 'chordates'. The chordates themselves split into the urochordates - things like sea squirts and other tunicates, the cephalochordates, where our little lancelet friend fits in, and the vertebrates - us.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The textbook view of these groups holds that echinoderms are the most 'primitive', or basal member of the deuterostomes. This is mainly due to the fact that they are unlike any other deuterostome - they are the odd ones out in terms of their symmetry, their locomotion, internal organisation et cetera. The next group up are the hemichordates, so called because they do have some of the characteristics which are normally associated with the chordates, in particular the presence of first signs of gill (pharyngeal) slits. The classical view argues that the urochordates (tunicates) are the most basal member of the chordate clade - they possess a hollow 'notochord' and dorsal nerve cord, but don't exhibit the body complexity, particularly body segmentation, seen in the remaining cephalochordates (lancelets) and vertebrates, which together comprise the Euchordata or 'true chordates'. This classical view is shown in the image below.

[Click to enlarge]
All of this is based on an apparent increase in complexity throughout the lineages - almost as if the principles behind that iconic 'ape to man' idea had been transplanted to explain the development from 'squishy' to 'us'. Those more like us are more advanced, those more squishy are, well, primitive. Of course, we're only dealing here with complexity in a morphological sense - the appearance of new and improved features on organisms over time. If we were to look at biochemical or intracellular physiological complexity, the earliest forms of life (bacteria) would kick our proverbial arse. This idea of a supposed correlation between being 'complex' and being 'advanced' isn't completely arrogant or conceited. The earliest forms of animal life at least were simple and squishy, and there is, overall, a general trend towards "complexity". But many squishy lineages developed incredible complexity - you only need to look at the cephalopods (octopuses, squid and cuttlefish) who are most closely related to snails and slugs, as well as the more 'complex' forms that have rediscovered the joys of a simple squishy lifestyle.
This view on the deuterostome phylogeny has been challenged before, with some success. However, the most significant recent change to the phylogeny really only focussed on the origins of the hemichordates (shown below), shifting them from being basal to the chordates, to being a more closely related sister taxon of the echinoderms. As you can see, that still maintains that nice, convenient transition from the more primitive and squishy sea squirts, through Amphioxus, up to us and the other vertebrates.
[Click to enlarge]
Not anymore.
A paper published in this last week's Nature throws some quite serious spanners into the classical works of deuterostome phylogeny. The researchers, Delsuc et al. (Delsuc interestingly held a post-doctoral position in Massey Univerity's Allan wilson Centre for Ecology and Evolution) used 146 common genes from 14 different deuterostomes to really hammer out the relationships between the groups. The results below are quite profound (ignore the black bits, just focus on the bottom coloured bits).
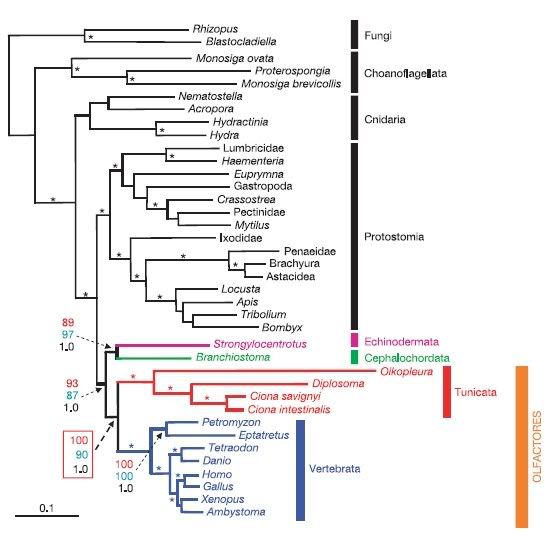
[Click to enlarge]
Essentially this provides two fundamental shifts:
Firstly, the urochordates are placed as the sister group to the Vertebrates, supporting a previously hypothesised clade called Olfactores. What this phylogeny implies is that we are more closely related to the sea squirt we are desperately trying to get rid of in Auckland Harbour than we are to Amphioxus. Now, this may not sound very important to some of you, but for anyone who has done even some basic evolutionary study, it means everything that we assume about the evolution and appearance of the vertebtrates, this gradual appearance of more 'vertebrate' features via the cephalochordates could quite possibly be absolute shite. Ipso facto, our (perhaps conceited) grouping "Euchordata" or "True chordates" could also be absolute shite.
Secondly, The cephalochordates are not only not directly basal to the vertebrates, they are actually placed as the sister group to the echinoderms in their own clade. Although the authors point out that this particular grouping doesn't have the statistical robustness as the urochordate-vertebrate grouping, it is nonetheless a rational interpretation of their data. This completely disrupts the "Chordata" group. From a systematics point of view, a lineage name applies to a common ancestor and all it's living and non-living descendants. Given that the chordates are now distributed all over the show (that is, they are polyphyletic) the term "chordate" may not, evolutionarily speaking, mean a hell of a lot. Unless of course we lowered the bar, if you will, and classified echinoderms (and by default hemichordates) as chordates. This naturally, is not likely to happen, because then basically we would be renaming all of the living deuterostomes as chordates. And that just won't do. No it won't. These two shifts can be seen in the simplified cladogram below.

[Click to enlarge]
This is the beauty of the tension that often arises between traditional morphological cladistics, or grouping organism relatedness based on overall similarity of body form and the sharing of structures, and that of molecular systematics - using genes to work out the same thing. Often they do come out with the same answer, but many times they don't. It's incredibly exciting when a study like this comes out, and challenges our assumptions about our 'knowledge' of morphological systems.
Perhaps more exciting than the actual repositioning of taxa within the deuterostome phylogeny is the implication for what this repositioning means for the evolutionary history of the deuterostomes themselves. The classical view suggests a steady aquisition and appearance of more 'vertebrate' like features through the lineages, with the cephalochordates representing the penultimate innovation before the appearance of the first vertebrate fish about 470 million years ago. Conversely, the echinoderms are seen (along with their sister group the hemichordates) as some bygone era of deuterstome evolution, a weird and wacky cousin that doesn't quite fit. Similarly, the urochordates, with their sessile habit and (apparently) simple body plan are seen as the primitive precursors to the euchordates.
According to the new study, however, both of these taxa can be viewed as highly advanced and derived lineages, which extensively modified the ancestral deuterostome body plan to become the forms that we see today. In the echinoderms an internal calcitic skeleton developed, along with radial symmetry and a particularly unique mode of locomotion, in the urochordates, the complete loss of the notochord in at least one sublineage, unique genetic simplicity and a lazy, squishy way of life.
And what of that ancestral body plan? The researchers suggest that the last common ancestor of the living deuterostomes may be far closer to Amphioxus than previously imagined - free living, bilateral symmetry, well developed metamerism (segmentation), distinct gill slits, and a pretty well organised brain and dorsal central nervous system. Essentially, all of these features that we assumed appeared just before we did may actually be the most ancestral, the most basic condition, Further those forms we assumed to be 'primitive' and distantly related are in fact extreme modifications of that basic body plan, bizarre yet equisite examples of evolution and adaptation, and in the case of the urochordates, closer to us in evolutionary time than we had ever imagined.
posted by Anonymous at 6:26 PM
![]()
<< Home